Highlights
Contents
Transplantation of firing pattern
Technical tools for SFEMG
Trigger
Delay
A few ways to get single muscle fibers in situ in man
Position of loudspeaker, remotely or closely, - GREAT
difference.
The birth of Macro EMG
Decrement in paralyzed rabbits, EAMG!!
Konzo
End of course
Transplantation of firing pattern
The Velocity Recovery Function (VRF) [1] i.e. dependence on muscle
propagation velocity (PV) on just preceding activity in the muscle fiber can be
studied in two ways
1. By double
stimulation with varying interpulse intervals
2. By mathematical
calculation of the relationship between the PV and the previous discharge
interval. The goodness of this calculation was tested as follows. PV was
measured from several hundred of discharges. The calculation of the VRF was
based on 200 discharges. The obtained algorithm was then applied to the
succeeding impulses to give the expected PV based on actual firing intervals.
This could be compared to the measured
PV values.
One way to see the similarity in VRF between different muscle fibers, a
number of them were stimulated with exactly the same impulse pattern. To obtain
this, we transplanted the firing pattern from a voluntary activation via tape
recorder and stimulator to the muscle and recorded PV in stimulated muscle
fibers.
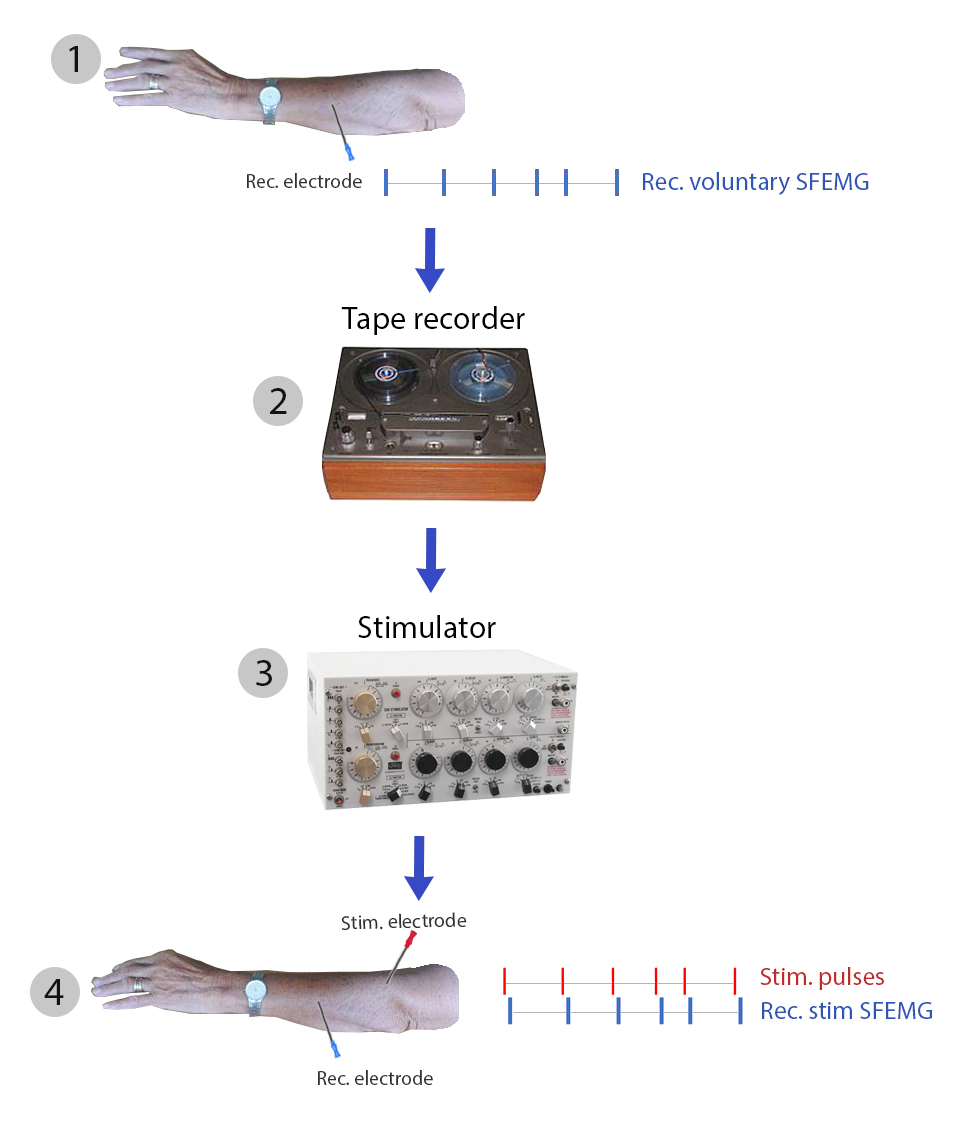
In practice: voluntary single muscle fiber action potentials obtained
during slight voluntary impulses (1) were stored on an analogue tape recorder
(2) . This pattern was then replayed to trigger a stimulator (3) connected to a
needle electrode that activated muscle fibers with the stored pattern (4).
SFEMG signals were recorded
The results of this is seen in the Figure.
Technical tools for SFEMG
During the development of the SFEMG method, a number of small but often
rather useful help tools or “gadgets” as some call them have been developed or at
least tested for use in clinical neurophysiology. They may have been more fun
than useful, but in some instances survived, even into commercially available
equipment.
Trigger
In order to see short duration signals, say 1 ms, that occur with a
frequency of 10/sec, i.e. they occupy 1% of the time, it is necessary to use a
high display sweep speed, often 02-0.5 ms/div. In more advanced oscilloscopes a
trigger was built in, i.e. the sweep started when the signal had reached an
amplitude level that could be optionally adjusted. This was not being used in
clinical neurophysiology at the time and we could not trigger on the SFAP. It
should be noted that the trigger function is present today in all EMG
equipment. The default is that all signals with an amplitude exceeding the
trigger level start the sweep. We sophisticated the trigger function further by
introducing an amplitude window, so that the sweep started as usual when the
amplitude was high enough, but was inhibited if the amplitude exceeded another
higher amplitude. The amplitude window could be adjusted optionally during the
recording. This made it possible to separate signals that had a lower amplitude
than the highest (Czekajewski, Ekstedt, Stålberg 1969). This feature has been
included in some EMG equipment, but I think that the possibilities it offers
have not yet been sufficiently recognized. Another problem was that only signal
segments coming after the time of trigger could be displayed on the sweep. This
can be seen in some of our early figures. This was solved by using a
delay-line, see below.
Delay
The problem with the missing part before the trigger had to be solved.
We tried to use a tape recorder with separate recording and reading heads. They
are separated by 1-2 cm and by using the correct tape speed; a delayed signal
should be obtained. This was not a practical solution - another type of delay
line was necessary. Our first solution was to use a one kilometer 14 paired
cable. We connected the ends and got a line of 28 km. This gave a delay of 93
µsec, sufficient to see some significant early parts of the signal. This later
became an electronic delay with resistors and condensers, and later a digital
delay, now the standard in all EMG equipment (Czekajewski, 1969).
A few ways to get single muscle
fibers in situ in man
During the development of SFEMG we needed to prove
that we were able to record from single muscle fibers. How to achieve this in
human?
Fibrillation potentials are usually considered to be generated by individual
fibers, so by recordings from such signals we could determine their
characteristics – good similarity to our SFEMG signals.
Another trick was to inject intramuscularly a small
dose of sodium citrate. This will chelate the sodium ions and fibers
start to fire independent of each other. A weak pain is felt.
Theoretically partial curarization should be a
method. At the initial stage of neuromuscular block of the voluntary activated
signal, individual building blocks (signals from single muscle fibers) can be
seen. When the jitter increased individual components in possible compound
signal started to show increased jitter and often increased latency. Thus
single fiiber action potential (a.p.) revealed themselves as single fiber a.p.
With further effect of the curare, these jittering components also show
intermittent and then complete block, The all-or-none behavior strongly
indicated that the spike was from a single muscle fiber. It is a very short
moment when this occurs and therefore this technique never became a practical
method to prove single fiber characteristics. Often however, the spiky signal
under exploration showed an all-or-none behavior upon curarization, i.e. was
present or absent. This was used as a strong indicator of a recording from a
single muscle fiber.
Position of loudspeaker, remotely or
closely, - GREAT difference.
In 1978 we gave an EMG course in Bombay
(Willison-England, Trojaborg -Denmark and me-Sweden). One evening we were
kindly invited to Mr Engineer’s for dinner. He had also invited Ravi Shankar,
relatively early in his career. All of us, around 30 persons, were seated
on the floor to listen to his music performed on sitar and some other
instruments played by Mr Shankar’s companions. Since the listening group
was large and the space quite big, a loudspeaker was placed at the end of the
room. Robin Willison was sitting close to this loudspeaker, perhaps 15 m away
from the players. A wonderful evening.

Next morning, Dr Willison had a comment on the sound
quality. He hear the muscle both directly, with the normal delay for sound in
air 50 msec for the 15 m, and from the loudspeaker, which produced the sound
without delay. His ear was very annoyed by this dual, non-simultaneous sound
input.
I well remember when they described the trick of using
different delays for the sound to loudspeakers in Westminster Abbey; those
speakers far away from the pulpit had a longer delay than those placed closer
to the pulpit.
The opposite situation occurred in our laboratory.
During a visit to us by Dr. Jasper Daube, he pointed out that a problem he had
with the EMG they used (at that time a Neuromatic 2000, the same as ours),
which gave an annoying time difference between the signal display and the sound
could not be detected in our lab.
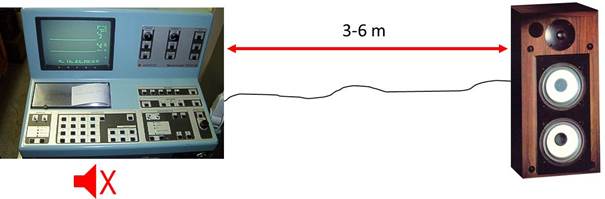
Fig showing an EMG equipment (Neuromatic 2000).
Internal loudspeakers are muted and separate loudspeakers 3-6 m away are
connected.
This EMG machine had a technical feature to first
record a signal segment for the total display time, say 20msec. Not until after
20 msec was the trace displayed on the screen, while the sound was “on line,”
in real time. With a very slow sweep, you could easily note this, but with
short sweep times only an expert like Daube could detect this. In our
situation, often using a sweep time of 20 msec, we had free standing
loudspeakers placed 3-6 m away (different for different rooms) instead of the
inbuilt speaker. Therefore the sound was delayed (time through air) by about
10-20 msec, throughout the range of sweep speeds that we used in routine. We
should have had a system by which the loudspeakers moved, depending on the
sweep speed!
The birth of Macro EMG
Together with Dr. Hilton-Brown, I was visiting
Columbus, Ohio in the late 70’ies. We discussed EMG (what else?) while walking
on the street. Intrigued by McComas’ relatively new publications on MUNE, we
asked if that method could be improved. His method was to use a metal strip as
the surface electrode over the muscle, usually small and thin muscles, such as
ADM or EDB. Our own studies had shown how motor units deep in a large muscle
such as biceps, did not give a signal to a surface electrode. Was there a way
to get a large metal electrode inside the muscle, close to the MU? Well, for
EMG with concentric needle electrodes (our standard) we always have a large
piece of metal inside the muscle – the cannula of the needle. So, on the street
in Columbus we decided to use the cannula for recording. Back home we made such
recordings with a SFEMG electrode, triggering on the SFEMG action potential and
averaging the time-locked cannula signal. We found that the amplitude depended
on the depth of the electrode. (By the way, this cannula signal is usually
subtracted from the tip-recorded signal in EMG. Since superficial positions of
the electrode give larger cannula signals, more will be subtracted in EMG, and
a superficial MU will be seen with a lower amplitude. We therefore modified the
SFEMG electrode by Teflon insulation except for the distal 15 mm, which gives a
large but standardized recording surface as long as the electrode is inserted
at least 15 mm. The triggering SFEMG electrode was placed 7.5 mm behind the
tip, in the center of the bare part of the cannula. This then became the
classic Macro EMG setup.
Decrement in paralyzed rabbits,
EAMG!!
Over the years I have experienced in a couple of important and
breath-taking moments.
One was the decrement and jitter in rabbits that had developed
antibodies to ACh-receptor. This was the first clue of postsynaptic receptor
involvement in MG (Heilbronn E, Mattsson C,
Stålberg E. Immune response in rabbits to a cholinergic receptor protein:
possibly a model for myasthenia gravis. Proc 3rd int cong on Muscle Diseases,
Newcastle 1974). The report came out just
after the report by Lennon VA, Lindstrom JM,
Seybold ME. Experimental autoimmune myasthenia: A model of myasthenia gravis in
rats and guinea pigs. J Exp Med 1975; 141:1365-1375
Konzo
A memorable moment in my scientific life was when in Oct 1991 we were asked
to examine two patients brought to Uppsala from the Congo. They had acute
spastic paraparesis and the pathophysiology was unknown. Insufficiently dried
cassava had produced cyanide poisoning with irreversible symptoms. They were
brought to Uppsala. For these patients, accompanied by family doctors and
some locals, this event must have been a shock. Flying in an airplane, coming
to a hospital, instruments giving electric shocks, and the noise in MRI. The
hospital staff did its best to understand their situation and help them adapt.
Blood chemistry - normal. Imaging techniques - normal. A full EDX
revealed no abnormalities in motor or sensory nerves. EEG was normal in one and
of low amplitude in the other patient. But when it came to transcortical magnetic
stimulation, we got the clue. No responses from arm or legs. The conclusion was
cortical inexcitability in this disease, which was called Konzo. The studies
continued and led to Dr Tshalas’ doctoral thesis. Later Dr Karin Eeg-Olofson
and eng PO Fällmar from Uppsala went to Zaire for field studies. Local
education on the cause was implemented.
End of course
This was the last day of an SFEMG course in Chapel
Hill NC, US in 1987 held in an anatomy auditorium. At the end of my good-bye
speech I tripped backwards and banged into the wall behind. This was assembled
by swinging segments about 2 m wide. Each segment could rotate around its
vertical middle axis. I pushed one segment which rotated 180 degree and I came
into a small pitch dark room. At the same time I heard laughter and applause. I
had really not said anything funny, but perhaps my exit was funny enough. When
I after a short moment return in front of the wall, I saw a skeleton, hanging
on the back of the segment, now widely exposed. The course really had a happy
end.
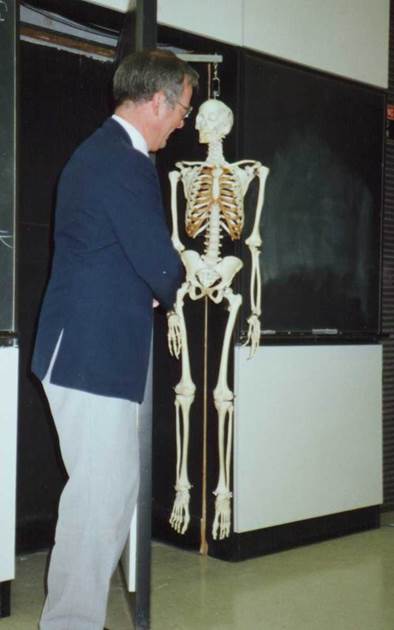
A most fantastic sortie, when Erik trips
Backwards through a rotating wall section
In the anatomy auditorium, disappears into
Darkness and a skeleton appears